Bálint Zsolt - Biró László Péter
A
lepkék színeváltozása
Bálint
Zsolt1 és Biró László
Péter2
1 Hungarian Natural History
Museum, 1088 Budapest, Baross utca 13, Hungary
2 Nanotechnology
Departement , Research Institute
for Technical Physics and Material Science , 1121 Budapest, Konkoly
Thege Miklós út 29-33, Hungary
A kolumbiai Andok entomológiai kutatásának úttörője, Anton Hermann Fassl (1876-1922) volt az, aki fölfigyelt a következő jelenségre: a magashegyi Loxurina-kardosboglár (Thecloxurina loxurina) hímjeinek szárnyfelszíne, amely rendszerint ragyogó mélykék színű, a tengerszint feletti magasság növekedésével elveszti tündöklő színét, és a szárnyon egyre inkább a vörös dominál. Közel száz év elmúltával bebizonyosodott, hogy a vörös foltokkal tarkított ibolyakék színű lepkék a rokon Atymna-kardosboglár-fajt képviselik (Thecloxurina atymna). Ennek ellenére a Fassl-féle megfigyelés részben mégiscsak helyes volt, mert nemcsak Kolumbiában, hanem Ecuadorban is a Loxurina azalacsonyabb, az Atymna pedig a magasabb tengerszint feletti magasságokban honos, bár egy szűk sávban elterjedésük fedi egymást. Az Atymna külső megjelenésében változékony: a faj magassági elterjedésének alsó régiójában teljesen ibolyakék példányok is előfordulnak, de minél feljebb haladunk, a szárnyfelületen a vörös rész egyre kiterjedtebb; és a 3800 m-en az Atymnávalegyütt megjelenik a teljesen vörös szárnyfelületű Atymnides-kardosboglár-testvérfaj is. A Loxurina ilyen magasságokban már nem fordul elő.
Hogyan lehetséges az, hogy a Loxurina testvérfajainak populációi (lásd "A színeváltós lepkék" 1-3. ábráját) a nagyobb magasságokban részben vagy teljesen elveszítik ragyogó kék színüket, és hogy a különböző színű egyedek csak bizonyos magasságokban fordulnak elő? Mi a válasz erre a jelenségre, és van-e jelentősége a fajképződés szempontjából?

A szerkezeti színek és szerepük
 A
lepkeidomú
rovarok teljes átalakulással fejlődnek, pete,
hernyó, báb és imágó (az
ivarérett lepke) egyedfejlődési alakjuk van. Az
imágó négy szárnyát zsindelyszerűen
egymásra boruló pikkelyek fedik (1. ábra).
Ez egyik fontos ismertető jegyük. Az egyedfejlődés
bábállapotának elején a pikkelyek,
még amolyan rügyecskeként, már ott sorakoznak
a szárnykezdemény lemezén. Maga a
színeződés és feltehetően ezzel együtt a
pikkelyszerkezetek kialakulása is a bábburokban,
néhány nappal az imágó kikelése
előtt zajlik. Ahogy azt számtalan kísérlet
bizonyítja, ilyenkor befolyásolható az egyed
szárnyainak színe és rajzolata.
A
lepkeidomú
rovarok teljes átalakulással fejlődnek, pete,
hernyó, báb és imágó (az
ivarérett lepke) egyedfejlődési alakjuk van. Az
imágó négy szárnyát zsindelyszerűen
egymásra boruló pikkelyek fedik (1. ábra).
Ez egyik fontos ismertető jegyük. Az egyedfejlődés
bábállapotának elején a pikkelyek,
még amolyan rügyecskeként, már ott sorakoznak
a szárnykezdemény lemezén. Maga a
színeződés és feltehetően ezzel együtt a
pikkelyszerkezetek kialakulása is a bábburokban,
néhány nappal az imágó kikelése
előtt zajlik. Ahogy azt számtalan kísérlet
bizonyítja, ilyenkor befolyásolható az egyed
szárnyainak színe és rajzolata.
Régóta ismert, de alaposan csak a közelmúltban leírt jelenség, hogy a lepkék ragyogó ibolya, kék és zöld színeit a pikkelyek különféle szerkezetei eredményezik. Az egyes pikkelyeken hosszanti irányban párhuzamosan végighúzódó, a pikkely lemezéhez pilléreken alulról támaszkodó gerinceket mélyen ülő keresztrudacskák kötik össze, cellácskákat alkotva. A cellácskák és a pikkely lemeze közötti teret nanoméretű szivacsos anyag tölti ki. Ezek kváziperiodikus elrendeződésük miatt fotonikuskristályhoz hasonlóan működnek, és ragyogó színt hoznak létre. Ez az egyik olyan pikkelyszerkezet, ami ragyogó színt képes létrehozni.
A legismertebb és részben bizonyított feltételezések szerint a szárnyakon látható szerkezeti színeknek az imágók ivari viselkedésében van szerepe. Kísérleti úton vizsgáltak a színpompás helikóniuszfajokat (Heliconius) szűk, egymást gyakran váltó, fényviszonyok szempontjából változatos esőerdei élőhelyeiken. A nőstény egyedek szerkezeti színei a szélsőséges fényhatások miatt a párválasztás során válogató hatást fejtenek ki. Egy másik feltételezés szerint a fényes szerkezeti szín a hímek közötti jeladásra szolgál: már távolról jelzi a rivális egyednek, hogy a terület foglalt, a "behatolás tilos". Ezt a feltételezést legújabban az ugyancsak esőerdei körülmények között élő morfók (Morpho) szerkezeti színeinek kialakulása kapcsán hozták fel magyarázatként amerikai kutatók. Egy további elmélet szerint a szerkezeti szín elvesztése a nőstényeken csak később fejlődött ki, a másodlagos ivari kiválasztódás hatására.
Robert Silberglied munkájának megjelenése óta a szerkezeti színek szerepét a lepkék esetében tehát kizárólag a vizuális kommunikációval és az ivari kiválasztódással magyarázták. Maga Silberglied volt az, aki ezt az irányt kijelölte, hiszen sorai szinte megkérdőjelezhetetlen, ex cathedra állításnak hatnak. Az általa és az ő nyomán kidolgozott elméletek egyike sem magyarázza, hogy rendszerint miért hiányzik a nem trópusi éghajlati viszonyok között tenyésző nőstények szárnyainak felszínéről a szerkezeti szín, vagy hogyan működik az ivari szelekció olyan fajok esetében, ahol a hím és a nőstény szárnyainak felszíne ugyanolyan színű.
A színváltás új elmélete
A kozmopolita boglárka (Polyommatus) genuszcsoportot vizsgálva fölfigyeltünk arra a jelenségre, hogy a nagy tengerszint feletti magasságokban vagy a magas szélességi körök alatt tenyésző fajok populációiban nemcsak a nőstény, hanem a hím is színt vált: elveszti szerkezeti kék színét, és megtévesztésig hasonló lesz a mérsékelt éghajlaton élő rendszerint barna nőstényhez. A jelenséget angolul diszkolorációnak neveztük el, amit magyarul úgy fordíthatnák, hogy a lepke színe "megváltozik", vagy hogy a lepke "színét váltja". A jelenségre új elméletet dolgoztunk ki. Eszerint a lepkék hideg vérű teremtmények, testük átmelegítéséhez és az izmok működtetéséhez szükséges hőenergiát sütkérezés útján nyerik, a következő módon: a nap felé fordítva a barna szárny a kéknél több hőenergiát képes begyűjteni, mivel az egész látható színképben elnyeli a sugárzást. A kék szín elvesztése a nőstények előtt már napokkal megjelenő hímek számára az életben maradáshoz előnyt jelent a kedvezőtlen éghajlati viszonyok között, olyan helyen, ahol kevesebb a napsütéses órák száma, mint például hegyvidéken vagy a magas északon.
Pásztázó elektronmikroszkóppal megvizsgáltunk és mérési kísérletek alá vetettünk egy testvérfajpárt, amelynek egyik tagja a kék színű, nagyrészt európai elterjedésű csipkés boglárka (Polyommatus daphnis), a másik pedig a színét váltott elbruszi Marcidusz-boglárka (Polyommatus marcidus) (lásd "A színeváltós lepkék" 10. és 11. ábráit). Bebizonyosodott, hogy a színét vesztett faj pikkelyeiből hiányoznak azok a szivacsos nanoszerkezetek, amelyek a testvérfaj kék színét adják. A mérési adatok azt is bebizonyították, hogy a színét váltott faj kék színű testvérénél (ugyanolyan körülmények között) másfélszer gyorsabban melegszik fel.
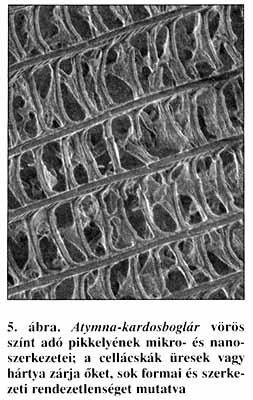
Eredményeink ismeretében pásztázó elektronmikroszkóp alatt megvizsgáltuk Fassl kolumbiai kardosboglárjait is. Vizsgálataink ugyanazt az eredményt mutatják, mint a már megvizsgált óvilági boglárka-testvérfajpár esetében. A ragyogó kék színű Loxurinában a szokványos mikroszerkezetek mellett megtalálható a pikkelyekben a nanoméretű szivacsosság (2. ábra) is. Ugyanez a nanoszerkezet az Atymna szárnyainak felszínén csak azokon a részeken figyelhető meg, ahol ibolyaszínt láthatunk (3. ábra). A többi vörös színű szárnyfelületen a pikkelyek cellácskái üresek (4. ábra), vagy erősen töredékes a szerkezetük (5. ábra). Az üres cellácskák nem működhetnek fotonikus kristályként. A fény teljes látható színképét elnyelik, és - feltehetően az óvilági fajpárhoz hasonlóképpen - a színét váltott ibolyásvörös Atymna és a vörös Atymnides ugyanannyi idő alatt a testébe több energiát képes koncentrálni, mint a kék Loxurina-testvérfaj.
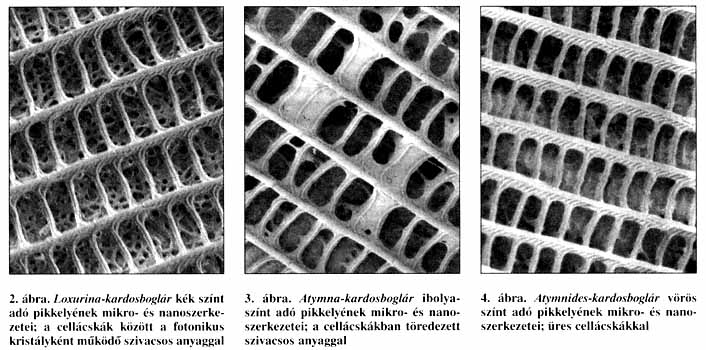
Nem ritka jelenség
Úgy tűnik, a lepkéknél hegyvidéken nem ritka a színeváltás jelensége. Például Peruban, ahol három faj szintén egyfajta magassági elrendeződést mutat: a Chachapoya kardosboglár (Thecloxurina chachapoya) teljesen kék színű hímje és az ezüstkék színű nősténye él legalacsonyab tengerszint feletti magasságban, fölötte a liláskék, szürkerózsás farkincás Ludovika-kardosboglár (Thecloxurina ludovica), majd a paramón, a magashegyi pusztán a vörös rozsdafarkú kardosboglár (Thecloxurina cillutincarae) (lásd "A színeváltós lepkék" 4-6. ábráit). Ez a csoport testvére a Loxurina-Atymna-Atymnides ágazatnak, de az Egyenlítőtől már jóval délebbre honos, magasabb szélességi körök mentén, más környezeti viszonyok között.
A jelenségre az északi féltekén is találunk bőven példát. A hispániai boglárka és a himalájai lángszinér (Heliophorus) színeváltós testvérfajpár, nem a kék vagy az ibolya, hanem a ragyogó zöld pikkelyek "színeváltását" szemléltetik (lásd "A színeváltós lepkék" 7-8-9. és 12. ábráit). A Fabresse-boglárka ("A színeváltós lepkék" 12. ábra) hispániai endemikus faj pikkelyei nanoszerkezet megváltoztatásával alkalmazkodtak a jégkorszak során oly gyakran változó környezeti viszonyokhoz. A délre szorult nem ezüstzöld Katalán-boglárka (lásd "A színeváltós lepkék" 9. ábráját) egy jégkorszak előtti, melegebb periódus maradványfaja lehet. Ugyanezek a folyamatok játszódhattak le a himalájai napszinér génusznál is (lásd "A színeváltós lepkék" 7-8. ábráját). Érdekes, hogy a tibeti faj színeváltása nem teljes: számos átmeneti alak származik a példányok lelőhelyéről, ezért feltételezzük, hogy a "diszlokáció" működésének mikéntje azon a populáción helyben vizsgálható lenne.
A szerkezeti színek szerepe
Dél-Amerikában a Fassl által vizsgált kardosboglárgenusz képviselői alapvetően más élőhelyeket népesítenek be, mint az esőerdei helikóniuszok és morfók: széles, egymást ritkán felváltó, fényeloszlás szempontjából egységes élőhelyeken honosak a lombkorona tetején (Loxurina), vagy a nyílt hegyi pusztán, az andesi paramón (Atymna és Atymnides). Meggyőzőbb azonban az - mérések útján általunk be is bizonyított magyarázat -, hogy a vizsgált magashegyi boglárka- és kardosboglárfajok esetében a pikkelyszerkezetek az egyed hőháztartásában játszanak fontos szerepet, és az általuk képzett színek a lepkék viselkedésében másodlagos jellegűek. Feltételezésünk szerint a Loxurina testvérfajai, akárcsak az elbruszi Marcidusz, azért vesztik el szerkezeti kék színüket, mert a nagyobb tengerszint feletti magasságokban a nem fotonikus kristályelv alapján működő pikkelyek az egyednek jobb energiafelhasználást biztosítanak.
A kardosboglárnál, akárcsak a példaként felhozott boglárka- és lángszinérfajok esetében, a szerkezeti szín szerepét magyarázó régi elméletnek ellentmondani látszik az, hogy éppen a legzordabb körülmények között élő, ezért minden eszközt bevetni kényszerülő magashegyi Atymnides veszti el az ivari kétalakúság eszközét. Ha az ivarok vizuális úton találják meg egymást, akkor miért éppen a mostoha viszonyok között élő fajoknál nehézkes az ivarok egymásra találása? Az általunk felállított és mérésekkel alátámasztott színeváltás-elmélet erre meggyőzőbb választ ad mind a nőstény, mind pedig a hím szempontjából.
A nőstény petéi a beérleléséhez szükséges energiamennyiséget üres mikro-cellácskákkal hamarabb be tudja juttatni testébe. Ez magyarázat arra, hogy a mérsékelt övi boglárkalepke-nőstények miért barnák. A trópusi éghajlat alatt élő nőstényeknek erre nincs szükségük, tehát ragyogó kékek. A Chachapoya és Loxurina vagy a csipkés boglárka nősténye ragyogó kék, és nincs szüksége erre a fortélyra, hiszen élete során több napenergiához jut, mint hegyvidéken élő vörös vagy barna társa, ahol a napsütéses órák száma jóval kevesebb. Joggal feltételezhető tehát, hogy a faj szempontjából a nőstény színváltása a fontosabb, ezért előbb alakul át a színe, mint a hímé.
A hímek kék szerkezeti színe, amely a népesség egyedsűrűségét hivatott szabályozni az egyedek versengő típusú hím-hím érintkezés rendkívüli gyakoriságával, olyan vidékeken tűnik el, ahol a faj számára a környezet már nem kedvező. A hímek agresszív versengő viselkedése zsebkendőnyi élőhelyen kipusztulást indikál. A meredek ökológiai lépcső mentén élő populáció, ahogy a kardosboglárfajok esetében is szemléltettük, a pikkelyszerkezet megváltoztatásával gyorsan alkalmazkodhat a megváltozott viszonyokhoz, és nemcsak a nőstények, hanem a hímek is akár egy pár generáció során is elveszíthetik szerkezeti színeiket. Ennek következményeként a populáció etológiája is gyökeresen átalakulhat, pl. a kék hímek "járőröző" viselkedése megváltozik, és a színeváltott egyedek területtartóvá válnak. A populáció egyedsűrűsége nagy mértékben növekedhet, ami nem elhanyagolható stratégia olyan helyeken, ahol minden egyes egyedre szükség van a faj túléléséhez.
Helyi fajképződés
A biológusok között még ma is vita tárgya: hogyan keletkeznek az új fajok? Többnyire elfogadott, hogy a párhuzamos vagy egymás mellett élő populációk tartós elszigetelődése új faj vagy fajok kialakulásához vezet. A helyi fajképződés elméletének, amikor is ugyanazon a helyen alakulnak ki a fajok, még számos ellenzője van. A párhuzamos és a mellérendelt fajképződés esetében már bizonyított a genetikai elkülönülést kiváltó mechanizmus módja. Ez a helyi fajképződés esetében nem ismert, vagy még nem mutatták ki meggyőző módon. A kutatások azonban tovább folynak. Legújabban az elméleti biológusok számítógépes modellezéssel próbálják szemléltetni, hogy a meredek környezeti lépcsők mentén valóban lehetséges a fajképződésnek ez a módja, mivel ezt a populáción belül az alkalmazkodási folyamatok biztosítják.
Feltételezzük, hogy a színeváltozás a helyi fajképződés egyik fontos eszköze. Ezt bizonyítják az általunk példaként felhozott boglárka-, lángszinér- és kardosboglár-testvérfajok. A lepkepopulációk környezeti változásokhoz való alkalmazkodásában az egyik leginkább működő mechanizmus az egyed pikkelyeinek nanoszerkezet-változása kell, hogy legyen. Ahogy a boglárkafajpáron végzett méréseink bizonyították, a különböző pikkely-nanoszerkezetű egyedek hőháztartása jelentősen eltér.
A legalacsonyabb és a legmagasabb pontokon kiegyenlített viszonyok uralkodnak: a faj számára a környezet a legkedvezőbb, illetve a legkedvezőtlenebb. Az itt élő alakok egyedei egyformák: csak kék Loxurina, illetve csak vörös Atymnides van, átmeneti alakok nincsenek. Pikkelyeik szerkezete rendezett (2-4. ábra). A két végpont között a helyzet átmeneti: ez nemcsak az Atymna-imágók változatos megjelenésen, az egyedek lila behintésének változatosságán, hanem a rendezetlen pikkelyszerkezeteikben is megnyilvánul (5. ábra). Az élőhelyen a kék Loxurina hímek az ezüstkék Loxurina nőstényekkel, míg az ibolyás-vöröses Atymna hímek az egyszínű vörös Atymna nőstényekkel párosodnak. A lejtő tetején már a teljesen vörös Atymnides tenyészik, ennek nősténye is vörösesbarna. Ha ez a helyzet nemzedékeken át állandósul és a különböző alakok az élőhely bizonyos mikrohabitatjaira specializálódnak, ez az állapot fajképződéshez vezet, hiszen ezek a formák elvesztik egymással kapcsolatukat, megindul a kiválasztódás, és a kék, a vörös és a lila populációk saját útjukra lépnek. A bemutatott boglárka-, lángszinér- és kardosboglárlepke példák azt mutatják, ez a fajképződési folyamat hamar lejátszódik, és nem egyszer éppen az ember az, aki az élőhelyeket feldarabolja, és ezeket a fiatal populációkat egymástól elszigeteli és önálló útra tereli. Van, amelyik képes az önálló életre és megtalálja a helyét a megváltozott viszonyok között, de olyan is - és sajnos egyre több - amely létezéséről már csak múzeumi példányok árulkodnak.
Kitekintés
Genetikailag
rögzül-e a "színeváltás"? Képes-e
újra ragyogó színbe öltözni és
ezáltal viselkedésmódot is váltatni egy
barnára vagy vörösre színeváltott
"populáció"? Izgalmas kérdések ezek. Ha
megtaláljuk a módját annak, hogyan
tenyésszünk üres
pikkelycellácskájú és szivacsos
nanoszerkezetű lepkéket ugyanabból a
populációból, akkor egy lépéssel
közelebb jutunk ahhoz a technikához, amely
segítségével az iparban is alkalmazható
nanoszerkezeteket tudunk előállítani olcsón
és nagy mennyiségben.
A munka az "Összehasonlító fiziomorfológiai és termodinamikai vizsgálatok boglárlepkefélék pikkelyein" című és T042972 számú OTKA kutatási program keretében készült.
| Természet Világa, | 135.
évfolyam, 7. szám, 2004. július http://www.chemonet.hu/TermVil/ http://www.kfki.hu/chemonet/TermVil/ |